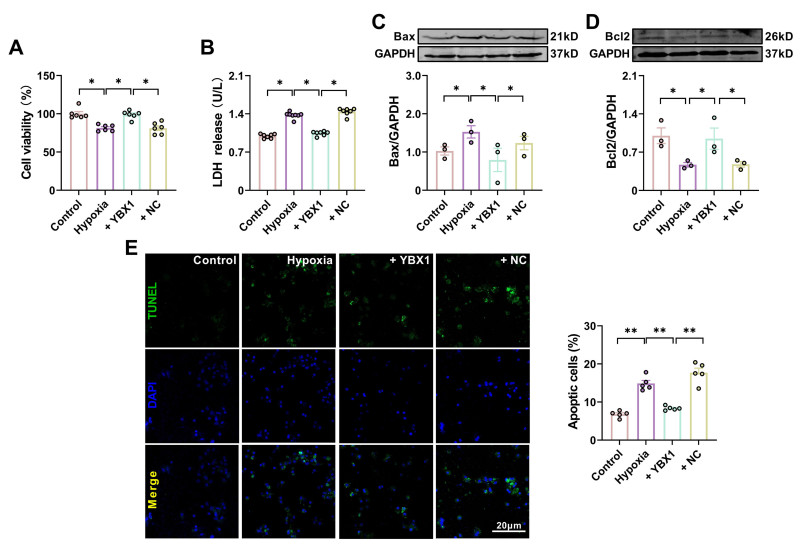
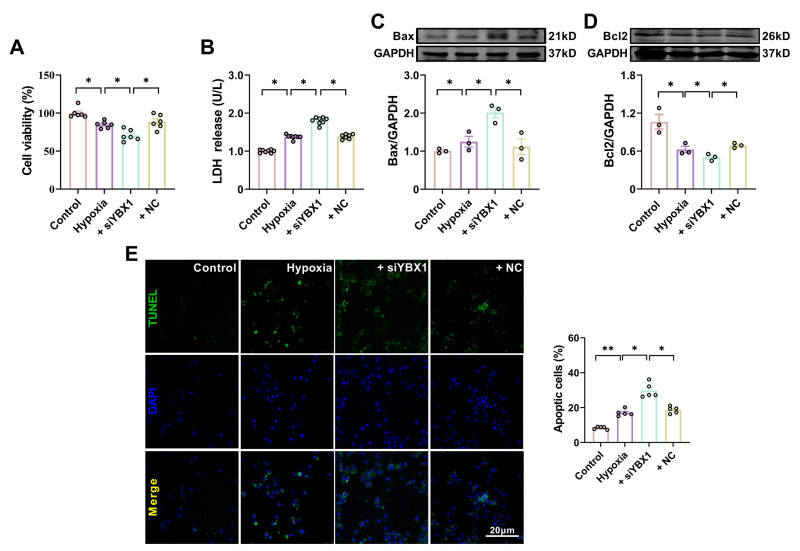
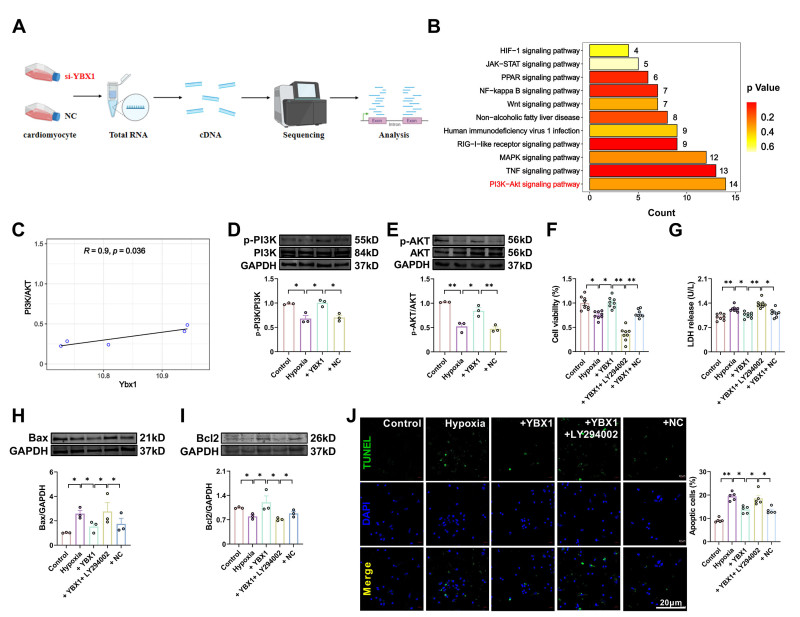
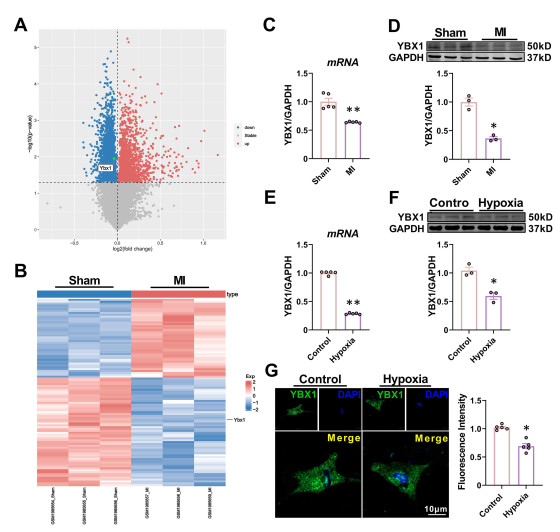
| Citation: | Fangfang Bi, Miao Cao, Yuquan Wang, Qingming Pan, Zehong Jing, Danyang Bing, Lifang Lyu, Tong Yu, Tianyu Li, Xuelian Li, Haihai Liang, Hongli Shan, Yuhong Zhou. YBX1 inhibits mitochondrial-mediated apoptosis in ischemic heart through the PI3K/AKT signaling pathway[J]. Frigid Zone Medicine, 2024, 4(1): 51-64. doi: 10.2478/fzm-2024-0006

|

| [1] |
Yuan X, Pan J, Wen L, et al. MiR-144-3p enhances cardiac fibrosis after myocardial infarction by targeting PTEN. Front Cell Dev Biol, 2019; 7: 249. doi: 10.3389/fcell.2019.00249
|
| [2] |
Acharya D. Predictors of outcomes in myocardial infarction and cardiogenic shock. Cardiol Rev, 2018; 26(5): 255-266. doi: 10.1097/CRD.0000000000000190
|
| [3] |
Bai X, Yang C, Jiao L, et al. LncRNA MIAT impairs cardiac contractile function by acting on mitochondrial translocator protein TSPO in a mouse model of myocardial infarction. Signal Transduct Target Ther, 2021; 6(1): 172. doi: 10.1038/s41392-021-00538-y
|
| [4] |
Luo Y, Zhang Y, Han X, et al. Akkermansia muciniphila prevents cold-related atrial fibrillation in rats by modulation of TMAO induced cardiac pyroptosis. EBioMedicine, 2022; 82: 104087. doi: 10.1016/j.ebiom.2022.104087
|
| [5] |
Lewington S, Li L, Sherliker P, et al. Seasonal variation in blood pressure and its relationship with outdoor temperature in 10 diverse regions of China: the China Kadoorie Biobank. J Hypertens, 2012; 30(7): 1383-1391. doi: 10.1097/HJH.0b013e32835465b5
|
| [6] |
Brennan P J, Greenberg G, Miall W E, et al. Seasonal variation in arterial blood pressure. Br Med J (Clin Res Ed), 1982; 285(6346): 919-923. doi: 10.1136/bmj.285.6346.919
|
| [7] |
Dong Y, Chen H, Gao J, et al. Molecular machinery and interplay of apoptosis and autophagy in coronary heart disease. J Mol Cell Cardiol, 2019; 136: 27-41. doi: 10.1016/j.yjmcc.2019.09.001
|
| [8] |
Elsässer A, Suzuki K, Lorenz-Meyer S, et al. The role of apoptosis in myocardial ischemia: a critical appraisal. Basic Res Cardiol, 2001; 96(3): 219-226. doi: 10.1007/s003950170052
|
| [9] |
Slee E A, Adrain C, Martin S J. Serial killers: ordering caspase activation events in apoptosis. Cell Death & Differentiation, 1999; 6(11): 1067-1074. doi: 10.1038/sj.cdd.4400601
|
| [10] |
Jose Corbalan J, Vatner D E, Vatner S F. Myocardial apoptosis in heart disease: does the emperor have clothes? Basic Res Cardiol, 2016; 111(3): 31.
|
| [11] |
Suresh P S, Tsutsumi R, Venkatesh T. YBX1 at the crossroads of non-coding transcriptome, exosomal, and cytoplasmic granular signaling. Eur J Cell Biol, 2018; 97(3): 163-167. doi: 10.1016/j.ejcb.2018.02.003
|
| [12] |
Cui Q, Wang C, Liu S, et al. YBX1 knockdown induces renal cell carcinoma cell apoptosis via Kindlin-2. Cell cycle (Georgetown, Tex.), 2021; 20(22): 2413-2427. doi: 10.1080/15384101.2021.1985771
|
| [13] |
Zhang H, Zheng W, Li D, et al. MiR-379-5p promotes chondrocyte proliferation via inhibition of PI3K/Akt Pathway by targeting YBX1 in osteoarthritis. Cartilage, 2022; 13(1): 19476035221074024. doi: 10.1177/19476035221074024
|
| [14] |
Hussain S A, Venkatesh T. YBX1/lncRNA SBF2-AS1 interaction regulates proliferation and tamoxifen sensitivity via PI3K/AKT/MTOR signaling in breast cancer cells. Molecular Biology Reports, 2023; 50(4): 3413-3428. doi: 10.1007/s11033-023-08308-5
|
| [15] |
Lin F, Zeng Z, Song Y, et al. YBX-1 mediated sorting of miR-133 into hypoxia/reoxygenation-induced EPC-derived exosomes to increase fibroblast angiogenesis and MEndoT. Stem Cell Res Ther, 2019; 10(1): 263. doi: 10.1186/s13287-019-1377-8
|
| [16] |
Huang S, Li X, Zheng H, et al. Loss of super-enhancer-regulated circRNA nfix induces cardiac regeneration after myocardial infarction in adult mice. Circulation, 2019; 139(25): 2857-2876. doi: 10.1161/CIRCULATIONAHA.118.038361
|
| [17] |
Pham T P, Bink D I, Stanicek L, et al. Long Non-coding RNA Aerrie Controls DNA Damage Repair via YBX1 to Maintain Endothelial Cell Function. Front Cell Dev Biol, 2020; 8: 619079. doi: 10.3389/fcell.2020.619079
|
| [18] |
Varma E, Burghaus J, Schwarzl T, et al. Translational control of Ybx1 expression regulates cardiac function in response to pressure overload in vivo. Basic Res Cardiol, 2023; 118(1): 25. doi: 10.1007/s00395-023-00996-1
|
| [19] |
Jing X, Yang F, Shao C, et al. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol Cancer, 2019; 18(1): 157. doi: 10.1186/s12943-019-1089-9
|
| [20] |
Zamaraev A V, Kopeina G S, Prokhorova E A, et al. Post-translational Modification of Caspases: The Other Side of Apoptosis Regulation. Trends Cell Biol, 2017; 27(5): 322-339. doi: 10.1016/j.tcb.2017.01.003
|
| [21] |
Ramachandra C J A, Hernandez-Resendiz S, Crespo-Avilan G E, et al. Mitochondria in acute myocardial infarction and cardioprotection. EBioMedicine, 2020; 57: 102884. doi: 10.1016/j.ebiom.2020.102884
|
| [22] |
Glaviano A, Foo A S C, Lam H Y, et al. PI3K/AKT/mTOR signaling transduction pathway and targeted therapies in cancer. Mol Cancer, 2023; 22(1): 138. doi: 10.1186/s12943-023-01827-6
|
| [23] |
Yang R, Li L, Hou Y, et al. Long non-coding RNA KCND1 protects hearts from hypertrophy by targeting YBX1. Cell Death Dis, 2023; 14(5): 344. doi: 10.1038/s41419-023-05852-7
|
| [24] |
Sepa-Kishi D M, Sotoudeh-Nia Y, Iqbal A, et al. Cold acclimation causes fiber type-specific responses in glucose and fat metabolism in rat skeletal muscles. Sci Rep, 2017; 7(1): 15430. doi: 10.1038/s41598-017-15842-3
|
| [25] |
Liu P, Yao R, Shi H, et al. Effects of cold-inducible RNA-binding Protein (CIRP) on liver glycolysis during acute cold exposure in C57BL/6 mice. Int J Mol Sci, 2019; 20(6): 1470. doi: 10.3390/ijms20061470
|
| [26] |
Xu B, Lang L M, Li S Z, et al. Cortisol excess-mediated mitochondrial damage induced hippocampal neuronal apoptosis in mice following cold exposure. Cells, 2019; 8(6): 612. doi: 10.3390/cells8060612
|
| [27] |
Kloetgen A, Duggimpudi S, Schuschel K, et al. YBX1 indirectly targets heterochromatin-repressed inflammatory response-related apoptosis genes through regulating CBX5 mRNA. Int J Mol Sci, 2020; 21(12): 4453. doi: 10.3390/ijms21124453
|
| [28] |
Su H, Fan G, Huang J, et al. YBX1 regulated by Runx3-miR-148a-3p axis facilitates non-small-cell lung cancer progression. Cell Signal, 2021; 85: 110049. doi: 10.1016/j.cellsig.2021.110049
|
| [29] |
David J J, Subramanian S V, Zhang A, et al. Y-box binding protein-1 implicated in translational control of fetal myocardial gene expression after cardiac transplant. Experimental Biology & Medicine, 2012; 237(5): 593-607. doi: 10.1258/ebm.2012.011137
|
| [30] |
Ma L, Singh J, Schekman R. Two RNA-binding proteins mediate the sorting of miR223 from mitochondria into exosomes. Elife, 2023; 12: e85878. doi: 10.7554/eLife.85878
|
| [31] |
Kishikawa T, Otsuka M, Yoshikawa T, et al. Satellite RNAs promote pancreatic oncogenic processes via the dysfunction of YBX1. Nat Commun, 2016; 7: 13006. doi: 10.1038/ncomms13006
|
| [32] |
Feng M, Xie X, Han G, et al. YBX1 is required for maintaining myeloid leukemia cell survival by regulating BCL2 stability in an m6A-dependent manner. Blood, 2021; 138(1): 71-85. doi: 10.1182/blood.2020009676
|
| [33] |
Chen J, Liu Z, Zhang H, et al. YBX1 promotes MSC osteogenic differentiation by activating the PI3K/AKT pathway. Curr Stem Cell Res Ther, 2023; 18(4): 513-521. doi: 10.2174/1574888X17666220805143833
|
| [34] |
Tuerxun T, Li X, Lou F, et al. YBX1 protects against apoptosis induced by oxygen-glucose deprivation/reoxygenation in PC12 cells via activation of the AKT/GSK3β pathway. Folia Biol (Praha), 2021; 67(4): 150-157. doi: 10.14712/fb2021067040150
|
| [35] |
Su W, Wang L, Zhao H, et al. LINC00857 interacting with YBX1 to regulate apoptosis and autophagy via MET and phosphor-AMPKa signaling. Mol Ther Nucleic Acids, 2020; 22: 1164-1175. doi: 10.1016/j.omtn.2020.10.025
|
| [36] |
Wang J, Hu K, Cai X, et al. Targeting PI3K/AKT signaling for treatment of idiopathic pulmonary fibrosis. Acta Pharm Sin B, 2022; 12(1): 18-32. doi: 10.1016/j.apsb.2021.07.023
|
| [37] |
Liu C, Chen K, Wang H, et al. Gastrin attenuates renal ischemia/ reperfusion injury by a PI3K/Akt/Bad-mediated anti-apoptosis signaling. Front Pharmacol, 2020; 11: 540479. doi: 10.3389/fphar.2020.540479
|
| [38] |
Feng C, Wan H, Zhang Y, et al. Neuroprotective effect of danhong injection on cerebral ischemia-reperfusion injury in rats by activation of the PI3K-Akt pathway. Front Pharmacol, 2020; 11: 298. doi: 10.3389/fphar.2020.00298
|
| [39] |
Li Y, Xia J, Jiang N, et al. Corin protects H(2)O(2)-induced apoptosis through PI3K/AKT and NF-κB pathway in cardiomyocytes. Biomed Pharmacother, 2018; 97: 594-599. doi: 10.1016/j.biopha.2017.10.090
|
| [40] |
Chen R, Chen T, Wang T, et al. Tongmai Yangxin pill reduces myocardial no-reflow by regulating apoptosis and activating PI3K/Akt/ eNOS pathway. J Ethnopharmacol, 2020; 261: 113069. doi: 10.1016/j.jep.2020.113069
|
 fzm-4-1-51_ESM.pdf
fzm-4-1-51_ESM.pdf
|

|
Figures(9)
Editorial Office: Frigid Zone Medicine, Editorial Office of Frigid Zone Medicine, Heilongjiang Health Development Research Center, Zhongshan Road 112, Xiangfang District, Harbin, 150036, China
Tel: 0451-87253028
E-mail: editorialoffice@frigidzonemedicine.com
Supported by:
Beijing Renhe Information Technology Co., Ltd.


 Submit
Submit


 DownLoad:
DownLoad: