 投稿系统
投稿系统

LCN2 aggravates diabetic cataracts by promoting ferroptosis in lens epithelial cells
doi: 10.1515/fzm-2024-0018
-
Abstract:
Background Cataracts are the leading cause of reversible blindness worldwide. Diabetic cataract (DC), a prevalent complication of diabetes mellitus, is characterized by its high occurrence, rapid progression, and severe impact. The prevalence of diabetes varies greatly between the northern and southern regions, with higher rates observed among northern residents. DC-induced lens opacity is mainly attributed to oxidative stress. However, it remains unclear whether ferroptosis, a form of regulated cell death, occurs in crystalline epithelial cells during the pathogenesis, which may represent a novel mechanism contributing to DC. Methods Transmission electron microscopy, quantitative assays for iron levels and reactive oxygen species (ROS), real-time quantitative polymerase chain reaction (RT-qPCR), western blotting, immunofluorescence, and immunohistochemistry were used to detect ferroptosis. Gene editing techniques were utilized to study the regulatory relationships among lipocalin 2 (LCN2), glutathione peroxidase 4 (GPX4), and ferritin heavy chain (FTH). Local knockdown of the LCN2 gene in B-3 cells and the eyes of Sprague Dawley (SD) rats was performed to verify and further explore the role and regulatory mechanisms of LCN2 in DC-associated ferroptosis. Results An in vitro model using high glucose levels and an in vivo model with streptozotocin-induced diabetes in SD rats were successfully established. Ferroptosis was observed in both in vitro and in vivo experiments. LCN2 protein was normally expressed in human and rat lens epithelial cells, but its expression significantly increased during ferroptosis. The ferroptosis inhibitor, ferrostatin-1 (Fer-1) effectively inhibited ferroptosis and reduced LCN2 protein expression. Notably, local knockdown of LCN2 via gene editing protected lens epithelial cells from ferroptosis in vitro and slowed the progression of DC in SD rats in vivo. Conclusion Our findings underscore the significant role of ferroptosis in the pathogenesis of DC, suggesting that selectively targeting LCN2 activation and enhancing ferroptosis resistance may offer a novel therapeutic approach for treating DC. -
Key words:
- lipocalin-2 /
- ferroptosis /
- diabetic cataracts
-

Figure 1. High glucose induces ferroptosis in B-3 cells in a dose-dependent manner
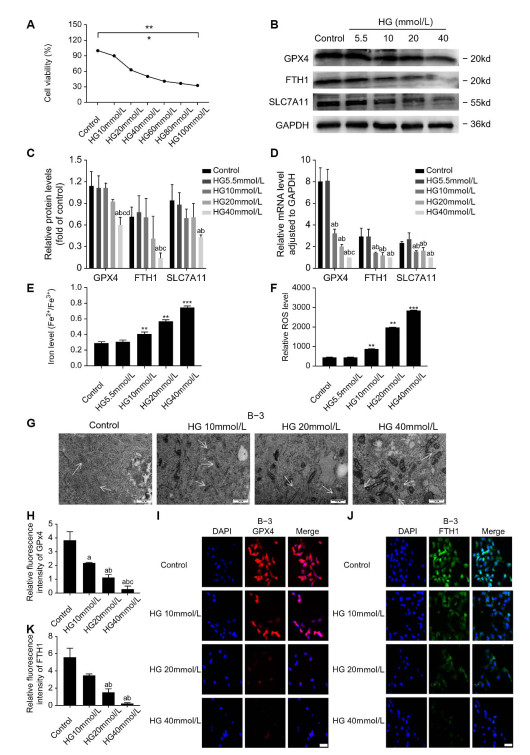
(A) To analyze the effect of high glucose concentrations on B-3 cell viability, CCK-8 assays were performed after treating the cells with culture media containing glucose at concentrations of 5.5, 10, 20, 40, 60, 80, and 100 mmol/L. The results are presented from three independent experiments. (B-D) The results of RT-PCR and Western blot analysis of GPX4, FTH1 and SLC7A11 when B-3 cells were treated with glucose concentration of 5.5 mmol/L and glucose concentration of 5.5 mmol/L + mannitol concentration of 40 mmol/L. Results of RT-PCR and Western blot analysis of GPX4, FTH1, and SLC7A11 when B-3 cells were treated with glucose at concentrations of 10, 20, or 40 mmol/L. The results are presented from three independent experiments. (a: Compared with the control group, b: compared with the 40 mmol/L mannitol group, c: compared with the 10 mmol/L high glucose group, d: compared with the 20 mmol/L high glucose group, all P < 0. 05) (E-F) Iron levels (Fe2+/Fe3+) and ROS levels in B-3 cells treated with glucose at concentrations of 5.5, 10, 20, or 40 mmol/L and glucose at a concentration of 5.5 mmol/L plus mannitol at a concentration of 40 mmol/L. The results are presented from three independent experiments. (**P < 0. 01; ***P < 0. 005) (G) When B-3 cells were treated with glucose at concentrations of 5.5, 10, 20, or 40 mmol/L, the ultrastructure of the cells was photographed by TEM. (H-K) When B-3 cells were treated with 5.5 mmol/L glucose and 5.5 mmol/L glucose + 40 mmol/L mannitol, the relative fluorescence intensity of GPX4 and FTH1 was measured by IF staining. When B-3 cells were treated with glucose at concentrations of 10, 20, or 40 mmol/L, the relative fluorescence intensities of GPX4 and FTH1 were measured by the IF staining method. The results are presented from three independent experiments. (a: Compared with the control group, b: compared with the 10 mmol/L high glucose group c: compared with the 20 mmol/L high glucose group, all P < 0. 05).

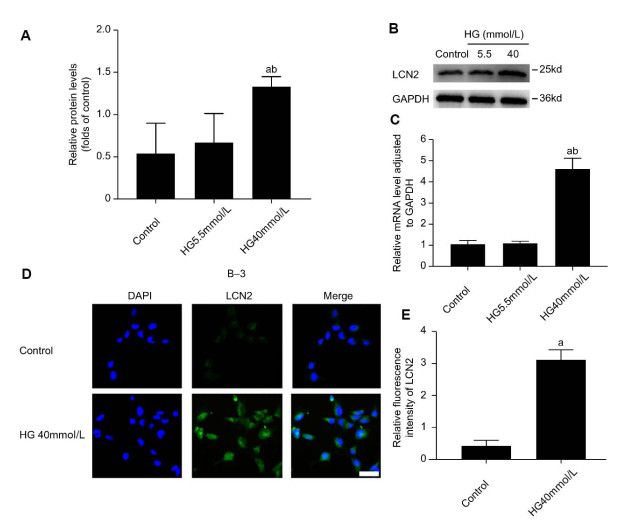
Figure 2. High glucose-induced differential expression of LCN2 in B-3 cells
(A-C) A blank control group and a high osmotic pressure control group were set up, and that result of RT-PCR and Western blot analysis of LCN2 are obtained when B-3 cells are treated with glucose concentration of 5.5 mmol/L, glucose concentration of 5.5 mmol/L + mannitol concentration of 40 mmol/L and glucose concentration of 40 mmol/L. The results are presented from three independent experiments. (D-E) A blank control group and a high osmotic pressure control group were set up. When B-3 cells were treated with glucose concentration of 5.5 mmol/L, glucose concentration of 5.5 mmol/L + mannitol concentration of 40 mmol/L and glucose concentration of 40 mmol/L, the relative fluorescence intensity of LCN2 was measured by if staining method. The results are presented from three independent experiments. (a: Compared with the control group, b: compared with the 40 mmol/L mannitol group, all P < 0. 05).

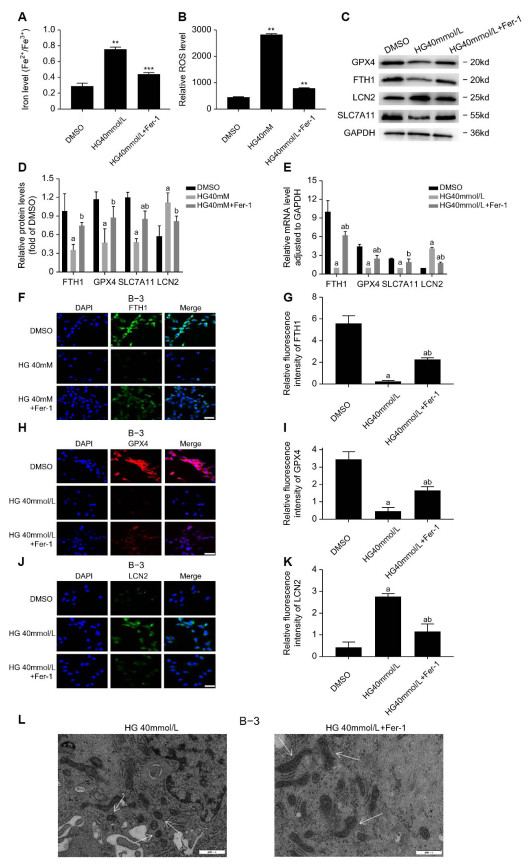
Figure 3. Fer-1 rescues high glucose-induced ferroptosis in B-3 cells
(A-B) Iron levels (Fe2+/Fe3+) and ROS levels in B-3 cells treated with DMSO, high glucose at a concentration of 40 mmol/L or Fer-1. The results are presented from three independent experiments. (C-E) Results of RT-PCR and Western blot analysis of GPX4, FTH1, LCN2 and SLC7A11 in B-3 cells treated with DMSO, high glucose at a concentration of 40 mmol/L, Fer-1. The results are presented from three independent experiments. (a: Compared with the DMSO group, b: compared with the 40 mmol/L high glucose group, all P < 0. 05) (F-K) When B-3 cells were treated with DMSO, 40 mmol/L high glucose or Fer-1, the relative fluorescence intensity of GPX4, FTH1, and LCN2 was measured by IF staining method. The results are presented from three independent experiments. (a: Compared with the DMSO group, b: compared with the 40 mmol/L high glucose group, all P < 0. 05). (L) When B-3 cells were treated with glucose at concentrations of 40 mmol/L or Fer-1, the ultrastructure of the cells was photographed by TEM.

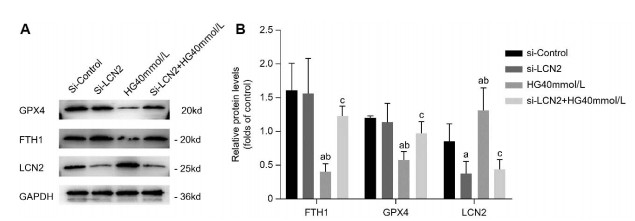
Figure 4. Silencing of LCN2 alleviates ferroptosis in B-3 cells
(A-B) Results of Western blot analysis of GPX4, FTH1 and LCN2 of B-3 cells for four groups of si-Control, si-LCN2, 40 mmol/L high glucose, and si-LCN2 + 40 mmol/L high glucose. The results are presented from three independent experiments. (a: Compared with the si-Control group, b: Compared with the si-LCN2 group, c: Compared with the 40 mmol/L high glucose group, all P < 0. 05).

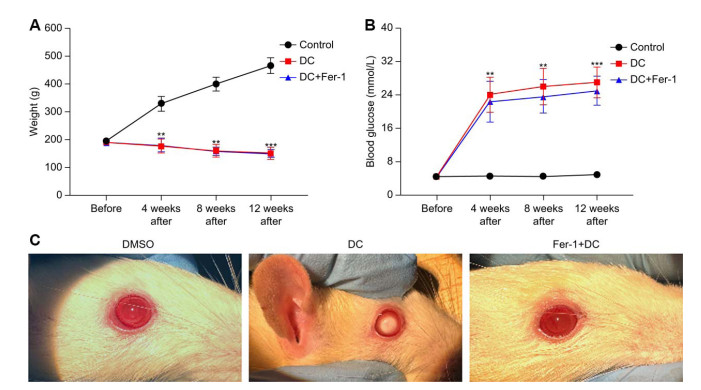
Figure 5. Development of DC model in SD rats
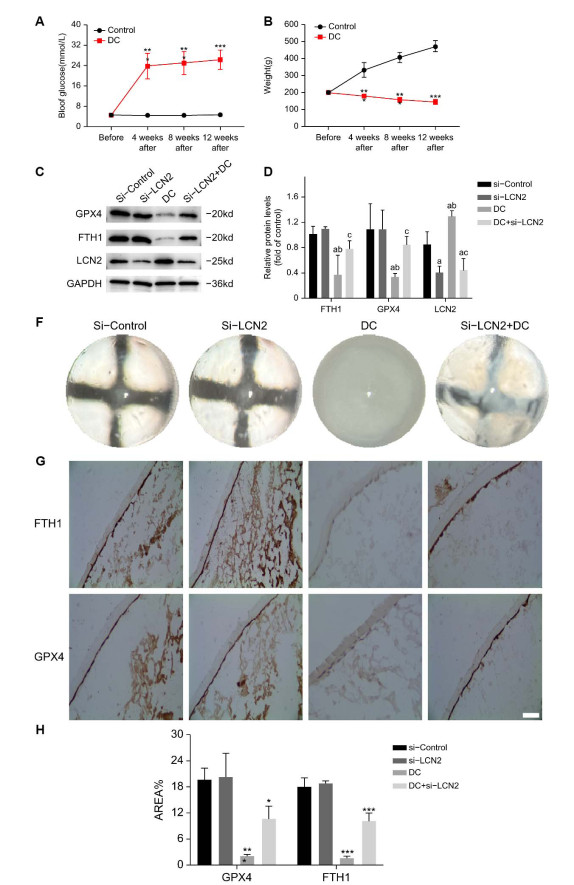
(A-B) The results of fasting blood glucose (mmol/L) and body weight (g) were measured before modeling, 4 weeks after modeling, 8 weeks after modeling and 12 weeks after modeling in the three groups of SD rats, they were DMSO group, DC group and Fer-1 treatment group. (**P < 0. 01; ***P < 0. 005). (C) The eye images of SD rats in the DMSO group, DC group and Fer-1 treatment group were taken after pupil dilation at 12 weeks after modeling.

Figure 6. Ferroptosis of lens epithelial cells isolated from SD rats with diabetes cataract
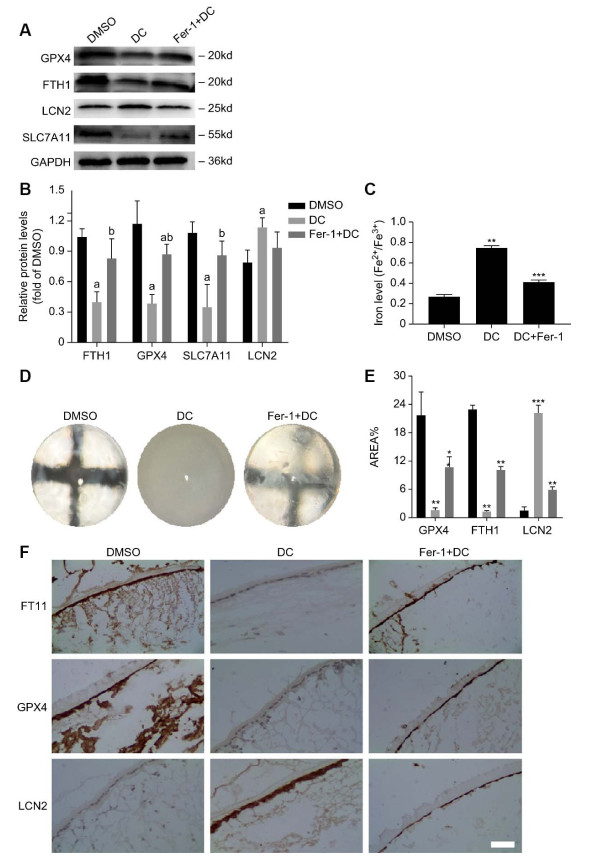
(A-B) They were three groups of SD rats (DMSO group, DC group and Fer-1 treatment group). The Western blot analysis results of GPX4, FTH1, LCN2, and SLC7A11 in the anterior capsule of their lenses. The results are presented from three independent experiments. (a: Compared with the DMSO group, b: : compared with the 40 mmol/L high glucose group, all P < 0. 05). (C) They were three groups of SD rats (DMSO group, DC group and Fer-1 treatment group). The results of iron levels (Fe2+/Fe3+) in the anterior capsule of their lenses. The results are presented from three independent experiments. (D) They were three groups of SD rats (DMSO group, DC group and Fer-1 treatment group). These are pictures of their complete lenses taken out. (E-F) They were three groups of SD rats (DMSO group, DC group and Fer-1 treatment group). The average optical density of GPX4, FTH1, and LCN2 in their anterior capsule was measured using the IHC method (compared to DMSO). The results are presented from three independent experiments. (*P < 0. 05; **P < 0. 01; ***P < 0. 005).

Figure 7. LCN2 silencing inhibits ferroptosis in lens epithelial cells and delays the occurrence and development of diabetes cataract in anterior segment of SD rats
(A-B) The results of fasting blood glucose (mmol/L) and body weight (g) measured before modeling, 4 weeks after modeling, 8 weeks after modeling, and 12 weeks after modeling in SD rats of the Control and DC groups. (C-D) Western blot analysis of GPX4, FTH1, and LCN2 in the anterior capsule membrane of SD rats in four groups: si-Control, si-LCN2, DC, and si-LCN2+DC. The results are presented from three independent experiments. (a: Compared with the si-Control group, b: Compared with the si-LCN2 group, c: Compared with the DC group, all P < 0. 05). (E) Complete lens images of four groups of SD rats: si Control, si-LCN2, DC, and si-LCN2+DC. (FG) The average optical density of GPX4 and FTH1 was measured using IHC method for the anterior capsule of SD rats in four groups: si-Control, si-LCN2, DC, and siLCN2+DC (compared to the si-Control group). The results are presented from three independent experiments. (*P < 0. 05; **P < 0. 01; ***P < 0. 005).
Table 1. All primer sequences
Primer Nucleotide sequence (5′ - 3′) GPX4 F-CGATACGCTGAGTGTGGTTT- R-CGGCGAACTCTTTGATCTCTT- FTH1 F-TACCTGAATGAGCAGGTGAAAG- R-GATATTCCGCCAAGCCAGAT- SLC7A11 F-GGTTGCCCTTTCCCTCTATTC- R-CCTGGGTTTCTTGTCCCATATAA- LCN2 F-CCAGGACAACCAATTCCAGGG- R-GTGGCATACATCTTTTGCGGG- GAPDH F-CTGGGCTACACTGAGCACC- R-AAGTGGTCGTTGAGGGCAATG- F, forward primer; R, reverse primer  下载: 导出CSV
下载: 导出CSV
Table 2. Comparison of weight and fasting blood glucose values of SD rats
Index Control (N = 20) DC (N = 20) DC+Fer-1 (N = 20) F P Weight (g) Before modeling 193.65±7.51 192.35±6. 04 190.7±4.51 1. 16 0. 321 4 weeks after modeling 330. 73 ±26.64b 177.48±25. 25a, b 180.75±24. 66a 235. 18 < 0. 001 8 weeks after modeling 400. 82 ±25.26b, c 159.94 ±23. 25a, b, c 155.71 ±17. 39a, b, c 797. 68 < 0. 001 12 weeks after modeling 465. 42 ±28.54b, c, d 151.47 ± 21. 5a, b, c, d 147.05 ± 17. 77a, b, c, d 1255. 35 < 0. 001 Blood glucose (mmol/L) Before modeling 4.50±0.36 4.51±0.42 4.48±0.35 0. 033 0. 968 4 weeks after modeling 4.71±0.36b 24.11 ± 4. 13ab 22. 46 ± 4.81a, b 172. 15 < 0. 001 8 weeks after modeling 4.53±0.72 26.13 ± 4. 3a, b, c 24. 72 ± 4.00a, b, c 249. 85 < 0. 001 12 weeks after modeling 5.06±0.42b, c, d 27.14 ± 3.63a, b, c 26. 13 ± 3.42a, b, c 372. 13 < 0. 001 aCompared with the control group; bCompared with before modeling; cCompared with 4 weeks after modeling; dCompared to 8 weeks after modeling; SD, Sprague Dawley; DC, diabetic cataract
下载: 导出CSV
Table 3. Comparison of weight and fasting blood glucose values of SD rats
Index Control (N = 10) DC (N = 10) t P Weight (g) Before modeling 196. 55 ± 7. 33 198.51 ± 6. 26 0. 642 0. 529 4 weeks after modeling 331. 68 ± 42. 47a 178.15 ± 11. 03a 11. 065 < 0. 001 8 weeks after modeling 405. 21 ± 29. 31a, b 156.92 ± 16. 56a 23. 323 < 0. 001 12 weeks after modeling 470. 93±32. 77a, b, c 143.12 ± 14. 14a, b, c 29. 048 < 0. 001 Blood glucose (mmol/L) Before modeling 4.64±0.37 4.68±0.54 0. 194 0. 848 4 weeks after modeling 4.55±0.41 23. 85 ± 5. 02a 12. 124 < 0. 001 8 weeks after modeling 4.61±0.46 25. 08 ± 4. 53a 14. 224 < 0. 001 12 weeks after modeling 4.74±0.34 26. 31 ± 3. 73a 18. 204 < 0. 001 aCompared with before modeling; bCompared with 4 weeks after modeling; cCompared with 8 weeks after modeling
下载: 导出CSV
-
[1] World Health Organization. Health Topics/Diabetes. https://www.who.int/health-topics/ . Accessed on May 27, 2021.[2] Liu Y C, Wilkins M, Kim T, et al. Cataracts. Lancet, 2017; 390(10094): 600-612. doi: 10.1016/S0140-6736(17)30544-5 [3] Thiagarajan R, Manikandan R. Antioxidants and cataract. Free Radic Res, 2013; 47(5): 337-345. doi: 10.3109/10715762.2013.777155 [4] Murtha T, Cavallerano J. The management of diabetic eye disease in the setting of cataract surgery. Curr Opin Ophthalmol, 2007; 18(1): 13-18. doi: 10.1097/ICU.0b013e32801129fc [5] Foster P J, Wong T Y, Machin D, et al. Risk factors for nuclear, cortical and posterior subcapsular cataracts in the Chinese population of Singapore: The tanjong pagar survey. Br J Ophthalmol, 2003; (9): 1112-1120. [6] Nirmalan P K, Robin A L, Katz J, et al. Risk factors for age related cataract in a rural population of southern India: The aravind comprehensive eye study. Br J Ophthalmol. 2004; 88(8): 989-994. doi: 10.1136/bjo.2003.038380 [7] Husain R, Tong L, Fong A, et al. Prevalence of cataract in rural Indonesia. Ophthalmology, 2005; 112(7): 1255-1262. doi: 10.1016/j.ophtha.2005.02.015 [8] Dandona L, Dandona R, Naduvilath T J, et al. Population-based assessment of the outcome of cataract surgery in an urban population in southern India. Am J Ophthalmol, 1999; 127(6): 650-658. doi: 10.1016/S0002-9394(99)00044-6 [9] Chen S J, Liu J H, Shih H C, et al. Prevalence and associated factors of lens opacities among Chinese type 2 diabetics in Kinmen, Taiwan. Acta Diabetol, 2008; 45(1): 7-13. doi: 10.1007/s00592-007-0012-9 [10] Starr M R, Mahr M A, Smith W M, et al. Outcomes of patients with active diabetic macular edema at the time of cataract surgery managed with intravitreal anti-vascular endothelial growth factor injections. Am J Ophthalmol, 2021; 229(9): 194-199. [11] Chen X, Yu C, Kang R, et al. Iron metabolism in ferroptosis. Front Cell Dev Biol, 2020; 8(10): 590226. [12] Dixon S J, Lemberg K M, Lamprecht M R, et al. Ferroptosis: an irondependent form of nonapoptotic cell death. Cell, 2012; 149(5): 1060-1072. doi: 10.1016/j.cell.2012.03.042 [13] Zhao Y, Pan B, Lv X, et al. Ferroptosis: roles and molecular mechanisms in diabetic cardiomyopathy. Front Endocrinol, 2023; 14(4): 1140644. [14] Huang Q, Tian H, Tian L, et al. Inhibiting Rev-erbα-mediated ferroptosis alleviates susceptibility to myocardial ischemia-reperfusion injury in type 2 diabetes. Free Radic Biol Med, 2023; 209(11): 135-150 [15] Wu Y, Chen Y. Research progress on ferroptosis in diabetic kidney disease. Front. Endocrinol, 2022; 29(9): 945976 [16] Abella V, Scotece M, Conde J, et al. The potential of lipocalin-2/NGAL as biomarker for inflammatory and metabolic diseases, Biomarkers. 2015; 20(8): 565-71. doi: 10.3109/1354750X.2015.1123354 [17] El Shahawy M S, Hemida M H, Abdel-Hafez H A, et al. Urinary neutrophil gelatinase-associated lipocalin as a marker for disease activity in lupus nephritis, Scand. J Clin Lab Invest, 2018; 78(4): 264-268. doi: 10.1080/00365513.2018.1449242 [18] Wells J M, Brummer R J, Derrien M, et al. Homeostasis of the gut barrier and potential biomarkers. Am J Physiol Gastrointest Liver Physiol, 2017; 312(3): G171-G193. doi: 10.1152/ajpgi.00048.2015 [19] Shao S, Fang H, Dang E, et al. Neutrophil extracellular traps promote inflammatory responses in psoriasis via activating epidermal TLR4/IL-36R crosstalk. Front Immunol, 2019; 10(4): 746. [20] Javaid H M A, Ko E, Joo E J, et al. TNFα-induced NLRP3 inflammasome mediates adipocyte dysfunction and activates macrophages through adipocyte-derived lipocalin 2. Metabolism, 2023; 142(5): 155527. [21] Asdaq S M B, Mannasaheb B A, Orfali R, et al. Antidiabetic and antioxidant potential of crocin in high-fat diet plus streptozotocin-induced type-2 diabetic rats. Int J Immunopathol Pharmacol, 2024; 38(11-12): 3946320231220178. [22] Qiao Y Y, Ji J L, Hou W L, et al. tRF3-IleAAT reduced extracellular matrix synthesis in diabetic kidney disease mice by targeting ZNF281 and inhibiting ferroptosis. Acta Pharmacol Sin, 2024; 45(5): 1032-1043. doi: 10.1038/s41401-024-01228-5 [23] El Maksoud A I A, Al-Karmalawy A A, ElEbeedy D, et al. Symbiotic antidiabetic effect of Lactobacillus casei and the bioactive extract of Cleome droserifolia (Forssk. ) Del. on mice with type 2 diabetes induced by alloxan. Chem. Biodivers, 2024; 21(1): e202301397. doi: 10.1002/cbdv.202301397 [24] Goyal S N, Reddy N M, Patil K R, et al. Challenges and issues with streptozotocin-induced diabetes - A clinically relevant animal model to understand the diabetes pathogenesis and evaluate therapeutics. Chem Biol Interact, 2016; 224(1): 49-63. [25] Conrad M, Pratt D A. The chemical basis of ferroptosis. Nat. Chem Biol, 2019; 15(12): 1137-1147. doi: 10.1038/s41589-019-0408-1 [26] Jiang X, Stockwell B R, Conrad M. Ferroptosis: Mechanisms, biology and role in disease, Nat. Rev. Mol Cell Biol. 2021; 22(4): 266-282. doi: 10.1038/s41580-020-00324-8 [27] Gao M, Monian P, Quadri N, et al. Glutaminolysis and transferrin regulate ferroptosis. Mol Cell, 2015; 59(2): 298-308. doi: 10.1016/j.molcel.2015.06.011 [28] Gao M, Monian P, Pan Q, et al. Ferroptosis is an autophagic cell death process. Cell Res, 2016; 26(9): 1021-1032. doi: 10.1038/cr.2016.95 [29] Gao M, Yi J, Zhu J, et al. Role of mitochondria in ferroptosis. Mol Cel, 2019; 73(2): 354-363. e3. doi: 10.1016/j.molcel.2018.10.042 [30] Hou W, Xie Y, Song X, et al. Autophagy promotes ferroptosis by degradation of ferritin. Autophagy, 2016; 12(8): 1425-1428. doi: 10.1080/15548627.2016.1187366 [31] Tan S, Schubert D, Maher P. Oxytosis: a novel form of programmed cell death, Curr. Top Med Chem, 2001; 1(6): 497-506. doi: 10.2174/1568026013394741 [32] Chen Y, Fang Z M, Yi X, et al. The interaction between ferroptosis and inflammatory signaling pathways. Cell Death Dis, 2023; 14(3): 205. doi: 10.1038/s41419-023-05716-0 [33] Li S, Lu S, Wang L, et al. Effects of amygdalin on ferroptosis and oxidative stress in diabetic retinopathy progression via the NRF2/ARE signaling pathway. Exp Eye Res, 2023; 234(9): 109569. [34] Devireddy L R, Gazin C, Zhu X, et al. A cell-surface receptor for lipocalin 24p3 selectively mediates apoptosis and iron uptake. Cell, 2005; 123(7): 1293-1305. doi: 10.1016/j.cell.2005.10.027 [35] Ellermann M, Arthur J C. Siderophore-mediated iron acquisition and modulation of host-bacterial interactions. Free Radic Biol Med, 2017; 105(4): 68-78. [36] Li D, Yan Sun W, Fu B, et al. Lipocalin-2-The myth of its expression and function. Basic Clin Pharmacol Toxicol. 2020; 127(2): 142-151. doi: 10.1111/bcpt.13332 [37] Goetz D H, Willie S T, Armen R S, et al. Ligand preference inferred from the structure of neutrophil gelatinase associated lipocalin. Biochemistry, 2000; 39(8): 1935-1941. doi: 10.1021/bi992215v [38] Xiao X, Yeoh B S, Vijay-Kumar M. Lipocalin 2: An emerging player in iron homeostasis and inflammation. Annu. Rev Nutr, 2017; 37(8): 103-130. [39] Dahl S L, Woodworth J S, Lerche C J, et al. Lipocalin-2 functions as inhibitor of innate resistance to Mycobacterium tuberculosis. Front Immunol, 2018; 9(11): 2717. [40] Guglani L, Gopal R, Rangel-Moreno J, et al. Lipocalin 2 regulates inflammation during pulmonary mycobacterial infections. PLoS One, 2012; 7(11): e50052. doi: 10.1371/journal.pone.0050052 [41] Saiga H, Nishimura J, Kuwata H, et al. Lipocalin 2-dependent inhibition of mycobacterial growth in alveolar epithelium. J Immunol, 2008; 181(12): 8521-8527. doi: 10.4049/jimmunol.181.12.8521 [42] Mei T, Wu J, Wu K, et al. Lipocalin 2 induces visual impairment by promoting ferroptosis in retinal ischemia-reperfusion injury. Ann Transl Med, 2023; 11(1): 3. doi: 10.21037/atm-22-3298 [43] Wu H, Santoni-Rugiu E, Ralfkiaer E, et al. Lipocalin 2 is protective against E. coli pneumonia, Respir Res, 2010; 11(1): 96. [44] Warszawska J M, Gawish R, Sharif O, et al. Lipocalin 2 deactivates macrophages and worsens pneumococcal pneumonia outcomes. J Clin Invest, 2013; 123(8): 3363-3372. doi: 10.1172/JCI67911 [45] Rahimi S, Roushandeh A M, Ahmadzadeh E, et al. Implication and role of neutrophil gelatinase-associated lipocalin in cancer: Lipocalin-2 as a potential novel emerging comprehensive therapeutic target for a variety of cancer types. Mol. Biol Rep, 2020; 47(1): 2327-2346. [46] Wang Y, Jia M, Yan X, et al. Increased neutrophil gelatinaseassociated lipocalin (NGAL) promotes airway remodelling in chronic obstructive pulmonary disease. Clin Sci (Lond), 2017; 131(11): 1147-1159. doi: 10.1042/CS20170096 [47] Ghosh S, Stepicheva N, Yazdankhah M, et al. The role of lipocalin-2 in age-related macular degeneration (AMD). Cell. Mol Life Sci, 2020; 77(5): 835-851. [48] Guardado S, Ojeda-Juárez D, Kaul M, et al. Comprehensive review of lipocalin 2-mediated effects in lung inflammation. Am J Physiol Lung Cell Mol Physiol, 2021; 321(4): L726-L733. [49] Tang W, Ma J, Gu R, et al. Light-induced lipocalin 2 facilitates cellular apoptosis by positively regulating reactive oxygen species/Bim signaling in retinal degeneration. Invest Ophthalmol Vis Sci, 2018; 59(12): 6014-6025. [50] Chen X, Kang R, Kroemer G, et al. Ferroptosis in infection, inflammation, and immunity. J Exp Med, 2021; 218(6): e20210518. [51] Chen Y, Yi X, Huo B, et al. BRD4770 functions as a novel ferroptosis inhibitor to protect against aortic dissection, Pharmacol Res, 2022; 177(5): 106122. [52] Tsurusaki S, Tsuchiya Y, Koumura T, et al. Hepatic ferroptosis plays an important role as the trigger for initiating inflammation in nonalcoholic steatohepatitis. Cell Death Dis, 2019; 10(6): 449. [53] Schnappauf O, Chae J J, Kastner D L, et al. The pyrin inflammasome in health and disease. Front. Immunol, 2019; 10(8): 1745. [54] Chauhan D, Vande Walle L, Lamkanfi M. Therapeutic modulation of inflammasome pathways. Immunol Rev, 2020; 297(1): 123-138. [55] Gupta U, Ghosh S, Wallace C T, et al. Increased LCN2 (lipocalin 2) in the RPE decreases autophagy and activates inflammasomeferroptosis processes in a mouse model of dry AMD. Autophagy, 2023; 19(1): 92-111. [56] Xie S S, Deng Y, Guo S L, et al. Endothelial cell ferroptosis mediates monocrotaline-induced pulmonary hypertension in rats by modulating NLRP3 inflammasome activation. Sci. Rep, 2022; 12(1): 3056. [57] Meihe L, Shan G, Minchao K, et al. The Ferroptosis-NLRP1 inflammasome: The vicious cycle of an adverse pregnancy. Front Cell Dev Biol, 2021; 9(8): 707959. [58] Luo L, Deng L, Chen Y, et al. Identification of lipocalin 2 as a ferroptosis-related key gene associated with hypoxic-ischemic brain damage via STAT3/NF-κB signaling pathway. Antioxidants (Basel), 2023; 12(1): 186. -

 点击查看大图
点击查看大图
计量
- 文章访问数: 652
- HTML全文浏览量: 364
- PDF下载量: 11
- 被引次数: 0

